


The Dysgenics of White Polygamy II
The Dysgenics of White Polygamy II
A follow-up examining the evidence for the assumptions presented in the first article


As I explained, the first article was a shallow and brief examination, on the assumption that the premises presented should be obvious. Evidently they are not. Many either refuse to admit or do not understand how (or why) the skewed genetic pool resulting from polygamy decreases intra-population genetic diversity. I will wind up repeating myself severally throughout this article, due to the basicness and similarity of the premises being denied. The simplest explanation is this: that the genetic drift which occurs over several generations creates enough genetic distance so as to normalize the rate of genetic divergence which occurs in monogamous procreation so as to maintain (or increase) effective population size. Any member of one sex who merges their lineage with multiple members of the opposite sex drives this rate away from the proportion of genetic drift which allows healthy reintegration after a few generations. The only way to offset this effect is to intermingle with other populations, which the goal ought to be to avoid. Otherwise genetic convergence compounds in each successive generation, which is at least slightly more propinquant than the last. This assumption is the one which is most in question, so I will use the assumption to explain certain ovserved failures, but also cover the reasons for this assumption. No piece of writing—let alone a short article—could feasibly give a comprehensive explanation of any of the principles being discussed, but there is certainly sufficient presence of these principles (and evidence for them) in academic examinations to lend mathematical credibility to what is, in my mind, an obvious assumption. I will move forward with the same assumption, as in the last article, though I will spend most of this article explaining the basis for said assumption.
The rate of consanguineous marriages in the Middle East cannot be entirely attributed to cultural mating choices of close relatives. It would be more accurate to say that, even if first cousin marriages were not preferentially treated, the Middle East would create a bottleneck anyways; so the Middle East’s accelerated preference towards consanguineous marriages simply gives us a glimpse of the inevitable. This is partially because the effect is inevitable, and partly because it is not isolated to the Middle East. Though the cursory graph I sketched up for the last article demonstrates the problem with polygamy in small communities, some people contested that a larger population size confounds this genetic convergence to such a degree that increased generational propinquity does not occur. However, high rates of consanguineous marriages occur in Africa, which generally does not share in the Middle East’s absence of an incest taboo; either in their Abrahamic age, or in indigenous African religions. Though it traditionally occurred in some places like Botswana (and even Ghana), almost exclusively the Muslim countries in Africa retain high rates of consanguineous marriages today. Marriage in Africa is traditionally exogamous. “The fundamental marriage rule followed throughout much of sub-Saharan Africa, and among the Luo, is very simple; no individual is allowed to marry someone from a related clan (including his own).”1 Granted, endogamous marriages are more traditional in the African countries which are today primarily Muslim. Whether this is because Islam has a retarding effect on incest taboos I am not interested in, because I don’t wish to spend any further time studying sub-Saharans; but I thought the anecdote was interesting enough to be considered.
The first necessary premise is effective population size (Ne). Ne is the ideal population size for heterozygosity’s decrease in proportion to the overall population, assuming the Hardy-Weinberg assumptions, which excludes inbreeding. So effective population size is determined under the assumption that no inbreeding is occurring. Therefore effective population size takes into account the increase of half-siblings from polygamous reproduction as non-viable potential partners. When the ratios of males and females mating is skewed from 1:1, effective population size (assuming no inbreeding) is depressed, because in each generation the amount of people who are half-related—and therefore excluded from each other’s potential mating pool—is increased. In other words, the mating pool of the average individual is significantly shrunk, unless inbreeding is permitted. This is further compounded by the increased fecundity of polygynous couples; so a meritocratically weighted allowance of wives such as proposed would cause even further depression of potential mates in larger sections of the populace. As men with more wives will generally produce more children, a larger section of the population will be skewed so as to be unable to mate with each other. The problem is not merely the increased propinquity among successive generations which directly result from polygamous reproduction, but also from the indirect depression of genetic diversity among potential mates in that generation.
It's true that most populations are large, but they don't necessarily act large. Thus, the rate of genetic drift is not really proportional to census population size (Nc). Rather, it's proportional to something more abstract — specifically, the effective population size (Ne). In an ideal population of sexually reproducing individuals, Ne will equal Nc. An "ideal" population has the following characteristics, and most deviations will decrease the effective population size:
1. There are equal numbers of males and females, all of whom are able to reproduce.
2. All individuals are equally likely to produce offspring, and the number of offspring that each produces varies no more than expected by chance.
3. Mating is random.
4. The number of breeding individuals is constant from one generation to the next.
As a note here, the requirement that there are equal numbers of males and females is in reference to the mating population; so once more, an application of eugenics to a monogamous society does not cause issues, because the ratio of reproductive participants remains monogamous (1:1), and therefore maintains the most efficient and beneficial level of genetic drift, such that similarly-descended individuals could remingle after several generations, with no effect on the diversity of lineages in the overall population.
Essentially, anything that increases the variance among individuals in reproductive success (above sampling variance) will reduce Ne (the size of an ideal population that experiences genetic drift at the rate of the population in question). For example, consider the effect of unequal numbers of mating males and females. In an ideal population, all males and all females would have an equal chance of mating. However, in situations in which one sex outnumbers the other, an individual's chance to mate is now affected by its sex, even if all individuals within each sex have an equal chance to mate.
…
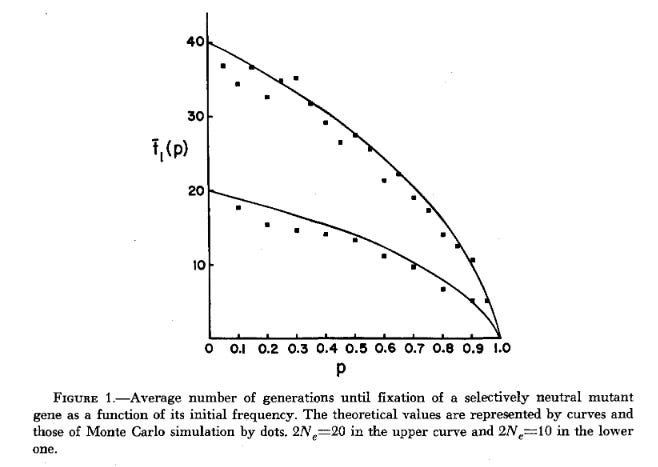
Therefore, fixation time scales with Ne. This time is maximized when p equals 0.5, and it falls off dramatically as one allele or the other becomes more rare at the generation we consider to be our starting point.2
So as I said in the first article, the issue is not with eugenic selection and exclusion—which in a monogamous system maintains the same proportion of genetic diversity in relation to genetic drift relative to the breeding population—but with disproportionate mating in any one direction.
This problem is not unprecedented. Mormons are a specimen of eugenics in spite of their polygamous history, not because of it - primarily because they are of good stock, and today practice monogamy. The central body of Mormons have not practiced polygamy for over 130 years; during its ~40 year tenure, only 30 percent of families were polygamous3. Despite this, the effects are still present today. More human disease genes have been discovered in the study of Utah genetics than anywhere else in the world. Some fundamentalist Mormon break-off sects still practice polygamy today, and the effects are as expected.
Short Creek gives us an example of a small population—which should be large enough to prevent genetic convergence—experiencing a genetic collapse due to polygamy. The population is close to 8,000. 3/4 of the residents are direct descendants of two men.4 Mormons are certainly not ignorant of their ancestry. They trace their genealogies religiously; they’re not accidentally marrying their cousins. One could argue that propinquity laws in the LDS are not stringent enough; but as in the Middle East, it is obvious that this is an accentuating and accelerating factor rather than a causal one. Such prohibitions would only stave off the inevitable. If genetic proximity begins to increase in each generation as potential mate pools are depressed for some or many individuals, then exogamy would be the only way to remedy this - clearly not a favorable solution. A (very) small number have argued that the effects of long-term incestous mating are worthwhile in exchange for an increased IQ; the debilitating diseases present in Short Creek speak against this. And the inevitability of fixation of hereditary diseases speak further against this5, especially since reproduction between close relations would guarantee that such genes—which are everywhere—present.
If the question turns to genetic engineering, then refer to my first article to at least glean my thoughts on the matter.
Some will inevitably ignore the basic principle which underlies all of this argument (that polygyny results in genetic convergence), and maintain that sufficient population size will negate the effects, despite the compounding effect of a skewed ratio. Humans are not the only case study however. Many animals appear “polygynous,” yes, but they are not really. They are polyamorous; the females mate with multiple males as well as the reverse. This certainly provides a confounding factor, because there is a greater diversity of lineages which are mixing. But aside from the fact that this solution is undesirable and disgusting—which I am sure most or all interlocutors would agree—it is not entirely sufficient in species which do not have sufficiently short maturation and gestation times. The problem is not merely the fact that polygyny causes a decrease in Ne (as discussed above), but that this decrease necessarily ends in genetic convergence (which is seemingly ignored). “Populations with small effective sizes are at risk for inbreeding depression and loss of adaptive potential. Variance in reproductive success is one of several factors reducing effective population size (Ne) below the actual population size (N).”6 The definition and structuring of Ne specifically relies on the genetic drift of the population, which as discussed above is necessarily altered by a skewing of female:male reproductive success ratio.
It is defined as the size of an idealized population of breeding adults that would experience the same rate of (1) loss of heterozygosity, (2) change in the average inbreeding coefficient, or (3) change in variance in allele frequency through genetic drift as the actual population (Futuyma, 1998, Frankham et al., 2002). As effective population size decreases, the rate of loss of allelic diversity via genetic drift increases. Two consequences of this loss of genetic diversity, reduced fitness through inbreeding depression and reduced response to sustained directional selection (“adaptive potential”), are thought to elevate extinction risk (Frankham et al., 2002, Frankham, 2005).
Mating success by sex has a relatively low impact on Ne, but it is an effect which does exist and causes an inevitable effect, which is (to repeat myself ad nauseam) compounded in each generation. Aside from the fact that selective pressures due to frequent death causes more progressive allelic change in the animal kingdom, it still cannot offset the convergence.
The effective size of a population is often much less than its actual size (N). Fluctuating population size, variance in reproductive success, and unequal sex ratios all reduce Ne (Wright, 1938, Frankham, 1995). Fluctuating population size has the largest effect and unequal sex ratios the least.
…
Polygyny is expected to produce higher variance in male reproductive success than alternative mating systems because variance will increase as fewer males monopolize more mates. The effect of polygyny on Ne has been examined both theoretically (Nunney, 1993, Engen et al., 2007) and in multiple empirical studies with a focus on mammals (Nunney and Elam, 1994, Storz et al., 2001, Storz et al., 2002).
To state the obvious and prevent quotemining, the variance discussed here is the variance in reproductive success; not genetic variance. The variance in reproductive success—or the skewing of male:female reproductive ratio—results in a decrease in effective population size necessarily because effective population size assumes no inbreeding. What is being said is that polygamous mating decreases effective population size because mating cannot move forward at the same level without inbreeding. If the population size shrinks sufficiently, then inbreeding has to occur regardless: otherwise extinction occurs. Extinction will occur regardless if Ne shrinks to such a degree that incest is required for reproduction, because congenital defects will destroy the species. The only way to prevent this is by introducing new blood, or mating exogamously; which is wholly undesirable.
First, Ne could be depressed because the species’ polygynous lek mating system, in which a relatively small proportion of males on leks perform most copulations (Wiley, 1973, Young et al., 2000), elevates the variance in male reproductive success. The potential for mating skew to decrease Ne in lekking grouse has been recognized previously (Bellinger et al., 2003, Johnson et al., 2003, Johnson et al., 2004, Bouzat and Johnson, 2004), but the magnitude of the effect has not been studied.
…
A novel prediction from the simulations is that Ne/_N_ should decline as population size increases. This effect arises because leks become larger as population size increases and, in larger leks, the variance in mating success is inflated because a smaller proportion of males mate and the number (though not the proportion) of matings by the most successful males increases. Although the predicted increase in the variance in male reproductive success with population size arises from a model of the dynamics of lek mating, this pattern has also been described in data from the polygynous, but non-lekking, red-winged blackbird (Weatherhead, 2005) and in flour beetles (Pray et al., 1996).
Even in a non-proportional increase in offspring, the effects of Ne depression occur, because each generation reduces the amount of potential partners available for random mating (which assumes no inbreeding).
As mentioned before: animals are very rarely polygynous, but rather polyamorous. But this is often insufficient to prevent genetic convergence in species with longer gestation and/or maturation periods, especially where intra-lineage polygyny occurs. In a meritocratic distribution of partners this would be further compounded by a naturally meritocratic intra-lineage polygyny. If one man’s wives are related to each other—for example two sisters who are chosen for the most eugenic male specimen because they both have a common eugenic inheritance—the compounding effect of genetic diversity depression is greatly heightened. There is a confluence of factors which all lead to detrimental effects of the proposed system. It is also greatly increased by re-mating, where the species is more polygynous than polyandrous; where female mates are loyal to a male mate. It is further increased by not practicing exogamy.
We show that re-mating and intralineage polygyny are associated with increased pairwise relatedness in the population and a rise in average inbreeding coefficients. In particular, the latter resulted from a correlation between male relatedness and rutting location, with related males being more likely to rut in proximity to one another.
…
Two aspects of mating systems have been revealed in polygynous mammals which, although extremely rarely reported, are potentially highly significant to the link between philopatry and co-ancestry/inbreeding. These are ‘mate fidelity’ (or ‘re-mating frequency’), females mating with the same male in one or more distinct breeding attempts (but without the establishment of a pair bond, that is, in contrast to the permanent pair bonds exhibited in monogamous species), and ‘intralineage polygyny’, whereby female relatives show a propensity to mate with the same male (Rossiter et al., 2005). Despite the clear potential for such behaviour in species with female philopatry, few studies report having capitalized upon new molecular techniques to test for such behaviour in polygynous systems, and so the extent to which it occurs is difficult to assess. However, there is evidence that it occurs in three polygynous mammals: red deer, grey seals and greater horseshoe bats. In red deer, Cervus elaphus, estimates of re-mating rates from behavioural observations of a small number of breeding hinds (29–32) over two consecutive 2-year periods ranged from 15.6% to 24.1% (Clutton-Brock et al., 1982b). In a study of paternity in grey seals, Halichoerus grypus, 30% of pups born to the same mother were found to be in full-sibs. In a species where litter size is usually one, and in which females mate in multiple years, this suggests many females were re-mating with the same male across years (Amos et al., 1995).7
This is especially true if we assume a eugenic reproduction model, where the primary eugenic male specimena continue to marry and mate with younger females, taking wives from successive generations to avoid the problems caused by aging female reproduction. I would assume that Mikka discounts the negative effects of aging male reproduction, but I could be incorrect here. If he does, then the concept is congregation of successive generations of females to one male mate so as to preserve eugenic female age to reproduction relationship. But this increases the average inter-relatedness of successive generations significantly. It’s a somewhat paradoxical interaction, as one might expect the children of more closely related mothers to return closer to the monogamous mean of effects on Ne. But in reality, this increases the amount of time and generations necessary for products of the same line (which has converged from several different lines) to begin mating again without procuring the consequences of inbreeding. But all the while, each successive generation becomes more closely related for the reasons above. So the most eugenic form of polygamy—that which would delay the effects of increasing propinquity for the maximal time—could not be purely meritocratic, as it requires an altered form of “random mating” in which multiple female partners must be unrelated to each other. But this leads to the more obvious dissemination of one patrilineal line into several matrilineal lines, which more intuitively causes depression of genetic diversity.
Rossiter et al. (2005) argue that increased pairwise relatedness is likely to strengthen ties between roosting females and therefore cooperation within social groups. In general, where generations of females are overlapping, intralineage polygyny and females re-mating with previous partners may raise co-ancestry but also increase the potential for inbreeding to occur (Chesser, 1991; Storz, 1999).
However this applies to any degree of relation in successive partners for one male. If Man1 takes Daughter1 of Man2 as a wife, and later takes the daughter of the same Man2’s Daughter2 as a second wife, the degree of interrelatedness causes the same effect, but to a slightly lesser degree than taking full sisters as wives. But to reiterate myself once more, the convergence of genetic interrelatedness in successive generations will necessitate such practices eventually. Though Short Creek is an extreme example, the convergence of every patrilineal line towards one or two common ancestors is necessitated by the skewing of mating ratios. As ancestry converges towards fewer and fewer patrilineal descents, the most recent common ancestor will also grow closer in this model.
https://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1224&context=bioscifacpub: all further citations until footnote 7 are from this source.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3546385/: all further citations are from this source.
